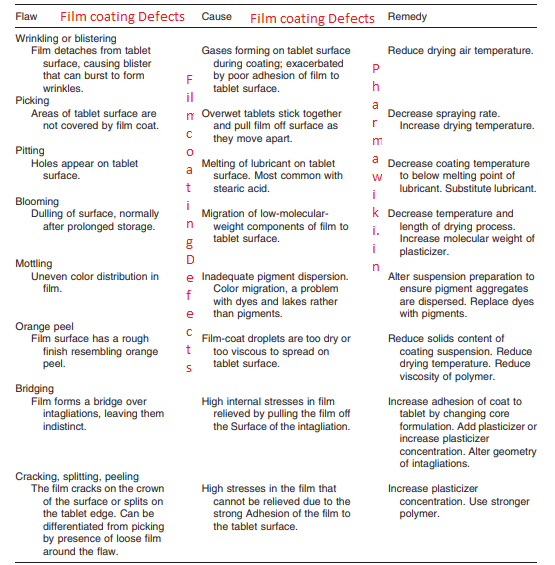
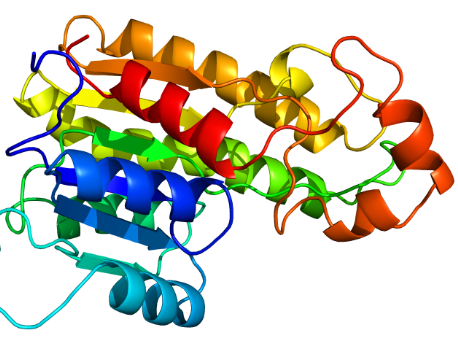
Protein Homology Modelling
Objectives
– Individual steps involved in calculating a protein homology model.
– Identify suitable templates for modelling.
– Outline the principles behind ab initio protein structure prediction.
– Describe the differences between homology modelling and ab initio structure prediction.
– Describe the major pitfalls in protein modelling.
Outline
§ Protein homology modelling
– Individual steps
– Caveats
– Pitfalls
§ Ab initio protein structure prediction
– Threading
– True ab initio methods
How Is It Possible?
§ The structure of a protein is uniquely determined by its amino acid sequence
(but sequence is sometimes not enough):
– prions
– pH, ions, cofactors, chaperones
§ Structure is conserved much longer than sequence in evolution.
– Structure > Function >> Sequence

How Often Can We Do It?
§ There are currently ~47000 structures in the PDB (but only ~4000 if you include only ones that are not more than 30% identical and have a resolution better than 3.0 Å).
§ An estimated 25% of all sequences can be modeled and structural information can be obtained for ~50%.
Worldwide Structural Genomics
§ Complete genomes
§ Signaling proteins
§ Disease-causing organisms
§ Model organisms
§ Membrane proteins
§ Protein-ligand interactions
Structural Genomics in North America
§ 10 year $600 million project initiated in 2000, funded largely by NIH.
§ AIM: structural information on 10000 unique proteins (now 4-6000), so far 1000 have been determined.
§ Improve current techniques to reduce time (from months to days) and cost (from $100.000 to $20.000/structure).
§ 9 research centers currently funded (2005), targets are from model and disease-causing organisms (a separate project on TB proteins).
Protein Homology Modelling PPT PDF for students researchers scholars
Homology Modeling for Structural Genomics
How Well Can We Do It?
How Is It Done?
§ Identify template(s) – initial alignment
§ Improve alignment
§ Backbone generation
§ Loop modelling
§ Side chains
§ Refinement
§ Validation ß
Template Identification
§ Search with sequence
– Blast
– Psi-Blast
– Fold recognition methods
§ Use biological information
§ Functional annotation in databases
§ Active site/motifs
Alignment
Improving the Alignment
Template Quality
§ Selecting the best template is crucial!
§ The best template may not be the one with the highest % id (best p-value…)
– Template 1: 93% id, 3.5 Å resolution L
– Template 2: 90% id, 1.5 Å resolution J
Evaluation of NMR Structures
What regions in the structure are most well-defined?
Template Quality – Ramachandran Plot
Backbone Generation
§ Generate the backbone coordinates from the template for the aligned regions.
§ Several programs can do this, most of the groups at CASP6 use Modeller:
http://salilab.org/modeller/modeller.html
Loop Modelling
§ Knowledge based:
– Searches PDB for fragments that match the sequence to be modelled (Levitt, Holm, Baker etc.).
§ Energy based:
– Uses an energy function to evaluate the quality of the loop and minimizes this function by Monte Carlo (sampling) or molecular dynamics (MD) techniques.
§ Combination
Loops – the Rosetta Method
§ Find fragments (10 per amino acid) with the same sequence and secondary structure profile as the query sequence.
§ Combine them using a Monte Carlo scheme to build the loop.
David Baker et al.
Side Chains
§ If the seq. ID is high, the networks of side chain contacts may be conserved, and keeping the side chain rotamers from the template may be better than predicting new ones.
Predicting Side Chain Conformations
§ Side chain rotamers are dependent on backbone conformation.
§ Most successful method in CASP6 was SCWRL by Dunbrack et al.:
– Graph-theory knowledge based method to solve the combinatorial problem of side chain modelling.
http://dunbrack.fccc.edu/SCWRL3.php
Side Chains – Accuracy
§ Prediction accuracy is high for buried residues, but much lower for surface residues
– Experimental reasons:
side chains at the surface are more flexible.
– Theoretical reasons:
much easier to handle hydrophobic packing in the core than the electrostatic interactions, including H-bonds to waters.
Refinement
§ Energy minimization
§ Molecular dynamics
– Big errors like atom clashes can be removed, but force fields are not perfect and small errors will also be introduced – keep minimization to a minimum or matters will only get worse.
Error Recovery
§ If errors are introduced in the model, they normally can NOT be recovered at a later step
– The alignment can not make up for a bad choice of template.
– Loop modeling can not make up for a poor alignment.
§ If errors are discovered, the step where they were introduced should be redone.
Validation
§ Most programs will get the bond lengths and angles right.
§ The Ramachandran plot of the model usually looks pretty much like the Ramachandran plot of the template (so select a high quality template).
§ Inside/outside distributions of polar and apolar residues can be useful.
§ Biological/biochemical data
– Active site residues
– Modification sites
– Interaction sites
Validation – ProQ Server
§ ProQ is a neural network based predictor that based on a number of structural features predicts the quality of a protein model.
§ ProQ is optimized to find correct models in contrast to other methods which are optimized to find native structures.
Structure Validation
§ ProCheck
http://www.biochem.ucl.ac.uk/~roman/procheck/procheck.html
§ WhatIf server
http://swift.cmbi.kun.nl/WIWWWI/
Homology Modelling Servers
§ Eva-CM performs continuous and automated analysis of comparative protein structure modeling servers
§ A current list of the best performing servers can be found at:
http://cubic.bioc.columbia.edu/eva/doc/intro_cm.html
Summary – Homology Modelling
§ Successful homology modelling depends on the following:
– Template quality
– Alignment (add biological information)
– Modelling program/procedure (use more than one)
§ Always validate your final model!
Fold Recognition and Ab Initio Protein Structure Prediction
Outline
§ Threading and pair potentials
§ Ab initio methods
§ Human intervention (what kind of knowledge can be used for alignment and selection of templates?)
§ Meta-servers (the principle, 3d jury)
§ Summary of take-home messages
Threading and Pair Potentials
§ Compares a given sequence against known structures (folds).
§ By using potentials that describe tendencies observed in known protein structures.
Potentials of Mean Force
Threading Methods Today
§ Problem: No protein is average
§ Interactions in proteins cannot only be described by pairs of amino acids
§ The information in the potentials is partly captured with sequence profiles
§ Today mostly used in HYBRID approaches in combination with profile-profile based methods
§ Potentials can be used to score models based on different templates or alignments
Ab Initio Methods
§ Aim is to find the fold of native protein by simulating the biological process of protein folding.
§ A VERY DIFFICULT task because a protein chain can fold into millions of different conformations.
§ Use it only when no detectable homologues are available.
§ Methods can also be useful for fold recognition in cases of extremely low homology (e.g. convergent evolution).
Fragment-based Ab Initio Modelling
§ Rosetta method of the Baker group:
– Submit sequence to a number of secondary structure predictors.
– Compare fragments of 3 and 9 residues to library from know structures.
– Link fragments together.
– Use energy minimization techniques (Monte Carlo optimization) to calculate tertiary structure.
Potentials for Finding Good Models
§ Use of energy potentials for scoring and computing models.
§ Potentials should make models more “native-like”.
§ These can be based on contact potentials, solvation potentials, Van der Waals repulsion and attractive forces, hydrogen bond potentials.
§ Globularity/radius of gyration (ab initio).
Problems with Empirical Potentials
Human Intervention
§ The best methods use maximum knowledge of query proteins.
§ Specialists can help to find a correct template and correct alignments.
Meta-Servers
§ Democratic modeling
– The highest score hit is often wrong.
– Many prediction methods have the correct fold among the top 10-20 hits.
– If many different prediction methods all have some fold among the top hits, this fold is probably correct.
Example of a Meta-Server
§ 3DJury http://bioinfo.pl/meta/
– Inspired by Ab initio modeling methods
• Average of frequently obtained low energy structures is often closer to the native structure than the lowest energy structure
– Find most abundant high scoring model in a list of prediction from several predictors
• Use output from a set of servers
• Superimpose all pairs of structures
• Similarity score based on # of Cα pairs within 3.5Å
– Similar methods developed by A. Elofsson (Pcons) and D. Fischer (3D shotgun).
3DJury
§ Because it is a meta-server it can be slow.
§ If queue is too long some servers are skipped.
§ Output is only Cα coordinates.
§ What to do with the rest of the structure?
§ Use e.g. maxsprout server to build sidechains and backbone atoms.
http://www.ebi.ac.uk/maxsprout/
Summary – Ab Initio Methods
§ Hybrid methods using both threading methods and profile-profile alignments are the best.
§ Use only Ab initio methods if necessary and know that the quality is really low!
§ Try to use as much knowledge as possible for alignment and template selections in difficult cases.
§ Use meta-servers when you can.
Protein Homology Modelling
Thomas Blicher
Center for Biological Sequence Analysis
Learning Objectives
After this lesson you should be able to:
– Explain the individual steps involved in calculating a protein homology model.
– Identify suitable templates for modelling.
– Outline the principles behind ab initio protein structure prediction.
– Describe the differences between homology modelling and ab initio structure prediction.
– Describe the major pitfalls in protein modelling.
Outline
§ Protein homology modelling
– Individual steps
– Caveats
– Pitfalls
§ Ab initio protein structure prediction
– Threading
– True ab initio methods
How Is It Possible?
§ The structure of a protein is uniquely determined by its amino acid sequence
(but sequence is sometimes not enough):
– prions
– pH, ions, cofactors, chaperones
§ Structure is conserved much longer than sequence in evolution.
– Structure > Function >> Sequence
How Often Can We Do It?
§ There are currently ~47000 structures in the PDB (but only ~4000 if you include only ones that are not more than 30% identical and have a resolution better than 3.0 Å).
§ An estimated 25% of all sequences can be modeled and structural information can be obtained for ~50%.
Worldwide Structural Genomics
§ Complete genomes
§ Signaling proteins
§ Disease-causing organisms
§ Model organisms
§ Membrane proteins
§ Protein-ligand interactions
Structural Genomics in North America
§ 10 year $600 million project initiated in 2000, funded largely by NIH.
§ AIM: structural information on 10000 unique proteins (now 4-6000), so far 1000 have been determined.
§ Improve current techniques to reduce time (from months to days) and cost (from $100.000 to $20.000/structure).
§ 9 research centers currently funded (2005), targets are from model and disease-causing organisms (a separate project on TB proteins).
Homology Modeling for Structural Genomics
How Well Can We Do It?
How Is It Done?
§ Identify template(s) – initial alignment
§ Improve alignment
§ Backbone generation
§ Loop modelling
§ Side chains
§ Refinement
§ Validation ß
Template Identification
§ Search with sequence
– Blast
– Psi-Blast
– Fold recognition methods
§ Use biological information
§ Functional annotation in databases
§ Active site/motifs
Alignment
Improving the Alignment
Template Quality
§ Selecting the best template is crucial!
§ The best template may not be the one with the highest % id (best p-value…)
– Template 1: 93% id, 3.5 Å resolution L
– Template 2: 90% id, 1.5 Å resolution J
Evaluation of NMR Structures
What regions in the structure are most well-defined?
Template Quality – Ramachandran Plot
Backbone Generation
§ Generate the backbone coordinates from the template for the aligned regions.
§ Several programs can do this, most of the groups at CASP6 use Modeller:
http://salilab.org/modeller/modeller.html
Loop Modelling
§ Knowledge based:
– Searches PDB for fragments that match the sequence to be modelled (Levitt, Holm, Baker etc.).
§ Energy based:
– Uses an energy function to evaluate the quality of the loop and minimizes this function by Monte Carlo (sampling) or molecular dynamics (MD) techniques.
§ Combination
Loops – the Rosetta Method
§ Find fragments (10 per amino acid) with the same sequence and secondary structure profile as the query sequence.
§ Combine them using a Monte Carlo scheme to build the loop.
David Baker et al.
Side Chains
§ If the seq. ID is high, the networks of side chain contacts may be conserved, and keeping the side chain rotamers from the template may be better than predicting new ones.
Predicting Side Chain Conformations
§ Side chain rotamers are dependent on backbone conformation.
§ Most successful method in CASP6 was SCWRL by Dunbrack et al.:
– Graph-theory knowledge based method to solve the combinatorial problem of side chain modelling.
http://dunbrack.fccc.edu/SCWRL3.php
Side Chains – Accuracy
§ Prediction accuracy is high for buried residues, but much lower for surface residues
– Experimental reasons:
side chains at the surface are more flexible.
– Theoretical reasons:
much easier to handle hydrophobic packing in the core than the electrostatic interactions, including H-bonds to waters.
Refinement
§ Energy minimization
§ Molecular dynamics
– Big errors like atom clashes can be removed, but force fields are not perfect and small errors will also be introduced – keep minimization to a minimum or matters will only get worse.
Error Recovery
§ If errors are introduced in the model, they normally can NOT be recovered at a later step
– The alignment can not make up for a bad choice of template.
– Loop modeling can not make up for a poor alignment.
§ If errors are discovered, the step where they were introduced should be redone.
Validation
§ Most programs will get the bond lengths and angles right.
§ The Ramachandran plot of the model usually looks pretty much like the Ramachandran plot of the template (so select a high quality template).
§ Inside/outside distributions of polar and apolar residues can be useful.
§ Biological/biochemical data
– Active site residues
– Modification sites
– Interaction sites
Validation – ProQ Server
§ ProQ is a neural network based predictor that based on a number of structural features predicts the quality of a protein model.
§ ProQ is optimized to find correct models in contrast to other methods which are optimized to find native structures.
Structure Validation
§ ProCheck
http://www.biochem.ucl.ac.uk/~roman/procheck/procheck.html
§ WhatIf server
http://swift.cmbi.kun.nl/WIWWWI/
Homology Modelling Servers
§ Eva-CM performs continuous and automated analysis of comparative protein structure modeling servers
§ A current list of the best performing servers can be found at:
http://cubic.bioc.columbia.edu/eva/doc/intro_cm.html
Summary – Homology Modelling
§ Successful homology modelling depends on the following:
– Template quality
– Alignment (add biological information)
– Modelling program/procedure (use more than one)
§ Always validate your final model!
Fold Recognition and Ab Initio Protein Structure Prediction
Outline
§ Threading and pair potentials
§ Ab initio methods
§ Human intervention (what kind of knowledge can be used for alignment and selection of templates?)
§ Meta-servers (the principle, 3d jury)
§ Summary of take-home messages
Threading and Pair Potentials
§ Compares a given sequence against known structures (folds).
§ By using potentials that describe tendencies observed in known protein structures.
Potentials of Mean Force
Threading Methods Today
§ Problem: No protein is average
§ Interactions in proteins cannot only be described by pairs of amino acids
§ The information in the potentials is partly captured with sequence profiles
§ Today mostly used in HYBRID approaches in combination with profile-profile based methods
§ Potentials can be used to score models based on different templates or alignments
Ab Initio Methods
§ Aim is to find the fold of native protein by simulating the biological process of protein folding.
§ A VERY DIFFICULT task because a protein chain can fold into millions of different conformations.
§ Use it only when no detectable homologues are available.
§ Methods can also be useful for fold recognition in cases of extremely low homology (e.g. convergent evolution).
Fragment-based Ab Initio Modelling
§ Rosetta method of the Baker group:
– Submit sequence to a number of secondary structure predictors.
– Compare fragments of 3 and 9 residues to library from know structures.
– Link fragments together.
– Use energy minimization techniques (Monte Carlo optimization) to calculate tertiary structure.
Potentials for Finding Good Models
§ Use of energy potentials for scoring and computing models.
§ Potentials should make models more “native-like”.
§ These can be based on contact potentials, solvation potentials, Van der Waals repulsion and attractive forces, hydrogen bond potentials.
§ Globularity/radius of gyration (ab initio).
Problems with Empirical Potentials
Human Intervention
§ The best methods use maximum knowledge of query proteins.
§ Specialists can help to find a correct template and correct alignments.
Meta-Servers
§ Democratic modeling
– The highest score hit is often wrong.
– Many prediction methods have the correct fold among the top 10-20 hits.
– If many different prediction methods all have some fold among the top hits, this fold is probably correct.
Example of a Meta-Server
§ 3DJury http://bioinfo.pl/meta/
– Inspired by Ab initio modeling methods
• Average of frequently obtained low energy structures is often closer to the native structure than the lowest energy structure
– Find most abundant high scoring model in a list of prediction from several predictors
• Use output from a set of servers
• Superimpose all pairs of structures
• Similarity score based on # of Cα pairs within 3.5Å
– Similar methods developed by A. Elofsson (Pcons) and D. Fischer (3D shotgun).
3DJury
§ Because it is a meta-server it can be slow.
§ If queue is too long some servers are skipped.
§ Output is only Cα coordinates.
§ What to do with the rest of the structure?
§ Use e.g. maxsprout server to build sidechains and backbone atoms.
http://www.ebi.ac.uk/maxsprout/
Summary – Ab Initio Methods
§ Hybrid methods using both threading methods and profile-profile alignments are the best.
§ Use only Ab initio methods if necessary and know that the quality is really low!
§ Try to use as much knowledge as possible for alignment and template selections in difficult cases.
§ Use meta-servers when you can.